Spike Timing Dependency Plasticity (STDP)
The Last Second
I'm not talking about the last second before I die. I have no knowledge of that experience and no way to report it when it happens. I'm talking about the last second you and I experienced while reading this very sentence. What facility in your brain gives this narrow less than one-second view of the world around you? Where is it physically located, and how does it work?
The search for this function is the sumation of my quest to understand what process in the brain allows us to participate in the moment.
I have a strong feeling that STDP is closely tied to the last second but after a long review of papers on the field, I find that the experts are as confused as I am, not a good sign for success. A good summary is by Feldman 'The Spike-Timing Dependence of Plasticity'.
It turns out that the basis for STDP is backpropagation (bAP) - that is, when the neuron spikes and sends its output down the axon chain, it also produces a second spike that propagates back into the dendrite tree. This seems to be what effectuates STDP in the synapses.
I found a way to modify the simulator to work with backpropagation and am moving forward with it. This solution involves sending Windows messages to various body parts during the simulation. You can see the progress on this solution in the Notes Volume 1 Chapter 5.
It is amateurs who have one big bright beautiful idea that they can never abandon. Professionals know that they have to produce theory after theory before they are likely to hit the jackpot.
-Francis Crick
My Theasus:
Without self-organization, the brain would start but soon stop - Locked down at an optimum condition. To keep the brain working, you need a little noise. Enough to jolt self-satisfied neurons out of their complacency and into action but not so much that the signal gets lost in the noise.
Aside from a little noise, you need some way that the brain can organize itself into a workable whole. It cannot be done by some sort of brain-within-brain composite brain that makes the final decisions based on inputs from all other parts of the brain. That duality requires that the 'inside brain' is made out of some stuff, not of this world.
I consider STDP to be the primary candidate for this self-organization property. I believe it is the primary Neural Correlate of Consciousness.
A good look at the field is presented in STDP an Overview [Markram]
Notes from Song
These notes were taken from the sentinel paper on STDP from Song, Mill and Abbott in March 2000 (available in the reference section)
- It must not have a global arbitrator involved. Local competition only
- Competition can also arise locally due to synaptic modification mechanisms that equilibrate at a per-set level of total synaptic innervation or activity [9 ]
- Spike timing provides a mechanism that can lead to competitive Hebbian learning without requiring global intracellular signaling, pre-set activity, or synaptic efficacy levels.
- Long-term strengthening of synapses occurs if presynaptic action potentials precede postsynaptic firing by no more than about 50 ms.
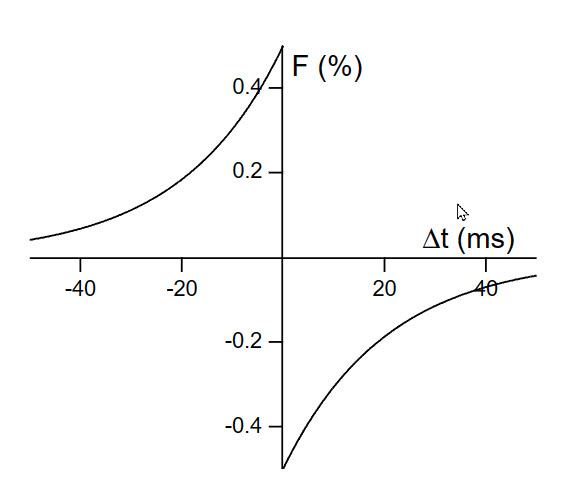
The STDP function plotted.
For example a spike comming in 20 mSec 'early' will improve that synapse by about 0.18 %. A 'late' incomming spike of 20 mSec will decrease that synapse by about -0.18%
The basic function used in this simulation:
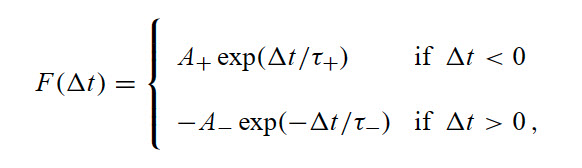
where τ+ and τ− determine the ranges of pre- to postsynaptic interspike intervals over
which synaptic strengthening and weakening occur. A+ and A− determine the maximum
amounts of synaptic modification, which occur when delta t is close to zero.
The temporal windows in the range of tens of milliseconds.
- The amplitude of synaptic modification, which is controlled by the parameters A+ and A−. In our simulations we use A− and A+ = 0.005
- In our modeling studies, we examine how STDP acts on the excitatory synapses driving an integrate-and-fire model neuron with N = 1000 excitatory and 200 inhibitory synapse.
- The excitatory synapses are activated by various types of spike trains: un-correlated spike trains generated by independent Poisson processes at various rates, bursts of action potentials with different latencies, and partially correlated spike trains. The model neuron also receives inhibitory input consisting of Poisson spike trains at a fixed rate of 10 Hz. In the simulations, excitatory synapses are modified on the basis of their pre- and postsynaptic spike timing, while inhibitory synapses are held fixed.
- STDP will strengthen short-latency excitatory inputs while weakening those with long latencies.
- Competition arises in a novel way, not due to a global signaling or growth factor, or to an artificially imposed balance of nonspecific synaptic decay and growth terms, but rather through a competition for control of the timing of postsynaptic action potentials
- As Hebb suggested 1 , synapses are only strengthened if their presynaptic action potentials precede, and thus could have contributed to, the firing of the postsynaptic neuron.
- Like any other Hebbian modification rule, STDP cannot strengthen synapses in the absence of postsynaptic firing. If for some reason the excitatory synapses to a neuron are too weak to make it fire, STDP cannot rescue them
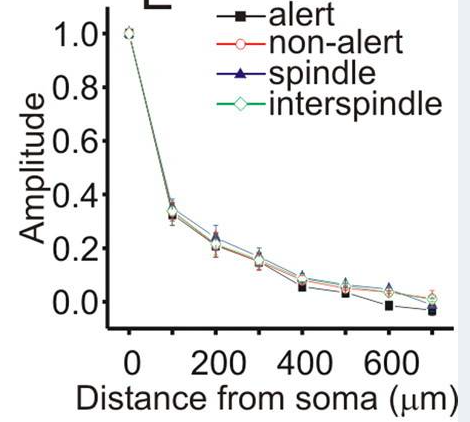
Backpropagation reduction over distance
Each time an AP fires it produces a bAP which travels backwards into the dendritic tree. The bAP disapates (unlike the AP) as it progresses. This curve shows the disapation rate [Bereshpolova Fig 5 E]